Master's Thesis Research
Statistical and Thermodynamical Properties of Lipid Membranes
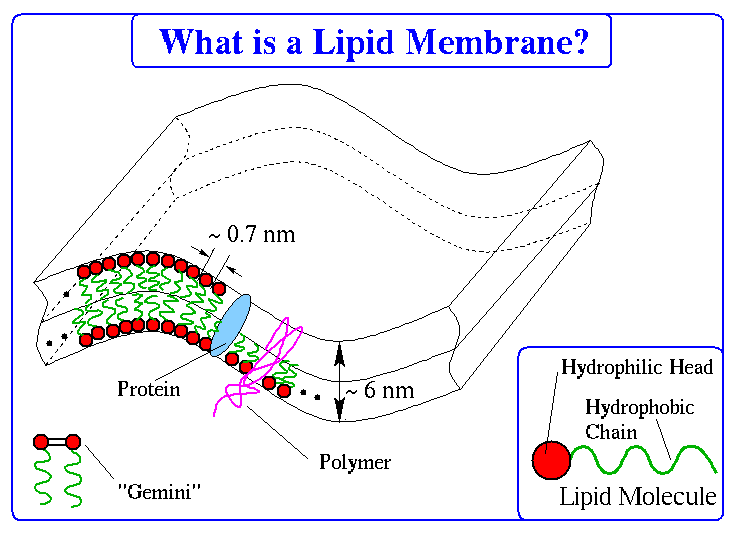
Membranes in biological systems such as cell walls usually consist of
fluid lipid bilayers, that is, two layers of tensid-like molecules with their
hydrophobic tails attached together and their hydrophilic heads directed
towards the solvent. Often, these lipid membranes form closed surfaces; little
blobs (lipid vesicles) of a size up to some microns.
The energies that bind adjacent lipid molecules (lipids) to each other
can range from 1...10^4 kT.
Therefore, some types of membranes are in constant
thermal motion and spontaneously undergo shape fluctuations, lipids are
free to move (flow) laterally.
However, due to
the fixed topology of the lipid molecules in the membrane, there is a finite
bending stiffness unlike i.e. in the fluid film of a soap bubble. Surface
tension is very small, on the other hand.
The membranes contain inclusions in their layers such as proteins or
long-chain polymers. Also, neighboring lipids can chemically bind and
thereby change the local stiffness (called "Gemini").
Cartoon of a Chunk of Lipid Membrane with Various Types of Inclusions
Besides analytical methods, numerical simulation has proved to be an
important means of investigating thermodynamical properties of these
systems. This includes Molecular Dynamics (MD) and
Monte Carlo (MC)
simulations. My Master
thesis is concerned with theoretical calculations and Monte Carlo simulation
of a lipid vesicle. Questions include thermal reduction of eff. stiffness,
spatial correlation lengths, and the determination of a persistence length.
Forces between particles embedded in the surface will be investigated.
 The following movie shows a
Monte Carlo (MC) simulation sequence of a lipid membrane vesicle represented
by 250 vertices (496 triangles). The movie shows an infinite loop
of ten frames. Five MC-sweeps were performed between successive frames.
(In a sweep, each vertex position is tried to be perturbed once)
The following movie shows a
Monte Carlo (MC) simulation sequence of a lipid membrane vesicle represented
by 250 vertices (496 triangles). The movie shows an infinite loop
of ten frames. Five MC-sweeps were performed between successive frames.
(In a sweep, each vertex position is tried to be perturbed once)
The expanding and shrinking bounding box shows width fluctuations of the
entire vesicle.
Click to get movie
Besides trying to move vertices, also the connection topology
of the triangulation is changed in a MC sweep. This assures "fluidity" of the
membranes, as it simulates the absence of shear stresses.
These "bond flips" rotate a link in the surrounding diamond that is
formed by the four adjacent links.
Here is another movie that shows
this process in detail right at the beginning of the simulation.
Here is the same at a later stage.
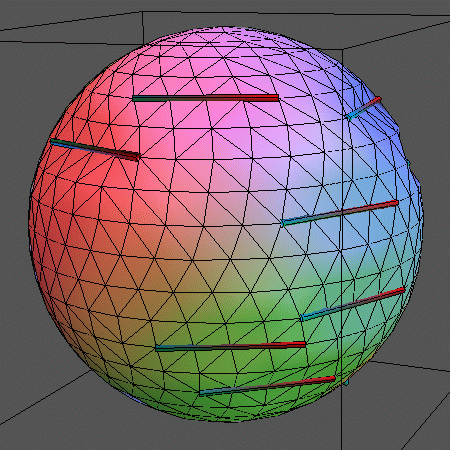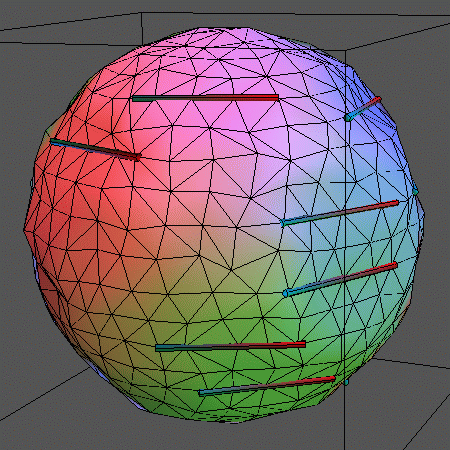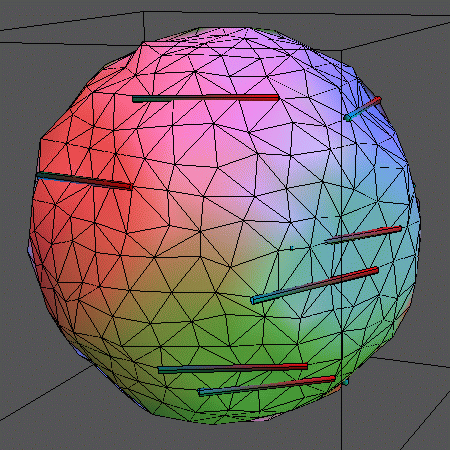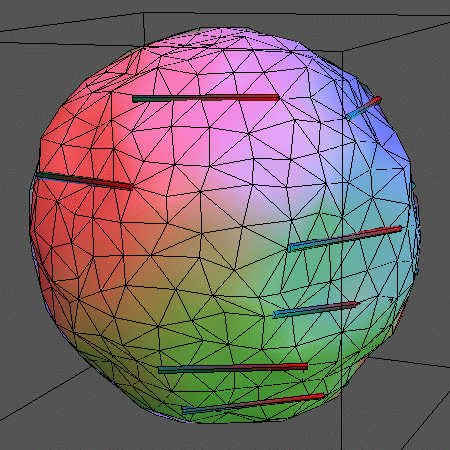
In addition to simulating the "naked" membrane, the surface can be
decorated by inclusions embedded in the lipids. This can be imagined like
logs of wood floating on water.
In the following animations, 14 rigid rods sit on the membrane represented by
500 vertices (996 triangles) and locally
constrain its movements by forcing 5 vertices each on a straight line. The
rods can leap between adjacent rows of vertices. Also, the above mentioned
vertex moves and bond flips are performed.
- Four frames in Animated Gif format,
270 kB, 450x450 pixels, 1.5 secs per frame
- MPEG-1 movie, 165 kB, 450x450 pixels,
16 Frames showing the membrane in steps of 1 MC-sweep, 25 frames/s (!)
- The same with 48 frames, 460 kB
- Same as the first MPEG, but in
MPEG-2 format, 359 kB
- Snapshot of the last frame, jpeg, 39 kB
Hint: To view MPEGs in Netscape, click on options..general
preferences..helpers. Then choose (or create) "video/x-mpeg" and as application
something like "mpeg_play %s > /dev/null 2>&1"
Get our publication on lipid membranes here.
Download my diploma thesis here (written in
English, PDF file, 750 kB).
Get my "Studienarbeit" (preparatory Master's thesis) in
uncompressed Postscript (220kB)
format or
gzipped (80 kB)
References:
R. Holzlöhner, M. Schoen:
"Attractive forces between anisotropic inclusions in the
membrane of a vesicle", Eur. Phys. J. B 12,
pp. 413-419 (1999)
J.H. Ipsen, C. Jeppesen:
"The Persistence Length in a Random Surface Model" , J. Phys. I France
5 (1995) 1563-71
Y. Kantor, D.R: Nelson:
"Crumpling Transition in Polymerized Membranes", Phys. Rev. Lett.
58 (1987) 2774-7
J.B. Fournier:
"Nontopological Saddle-Splay and Curvature Instabilities from
Anisotropic Membrane Inclusions" , Phys. Rev. Lett.
76 (1996) 4436-4439
D.R. Nelson, T. Piran, S. Weinberg et al.:
"Statistical Mechanics of Membranes and Surfaces",
Academic Press, London 1992
R. Golestanian:
"Fluctuation-Induced Interactions between Rods on a Membrane",
Phys. Rev. E 54 (1996) 6725-34
T.C Lubenski, F.C. MacKintosh:
"Theory of Ripple Phases of Lipid Bilayers",
Phys. Rev. Lett. 71 (1993) 1565 ff.
R. E. Goldstein, S. Leibler:
"Structural Phase Transitions of Interacting
Membranes" , Phys. Rev. A 40 (1989) 1025 ff.
E-mail: ronald.holzloehner @ web.de (remove spaces)

Last Modified: June 25, 2002
{kind=link}
{kind=link}
{kind=link}
{kind=link}